AMPHIPOLS: Properties of membrane protein/amphipol complexes
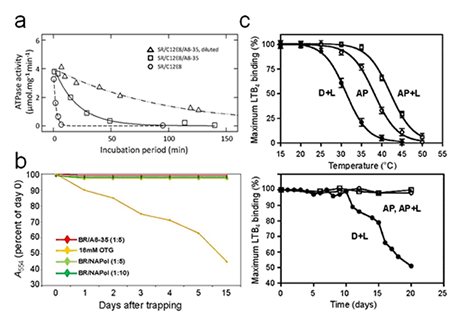
When properly prepared, MP/A8-35 complexes are essentially monodisperse (9, 10, 12). It is however difficult to totally avoid the presence of minor fractions of small oligomers, which can seriously complicate, in particular, radiation scattering experiments (9). Among the factors that can lead to aggregation are the use of too little APol at the trapping step (12), lowering the pH at or below neutrality (9), the presence of calcium ions (10), drifts from the nominal composition of the polymer (9, 10), and removal of the extra, free polymer that co-exists with MP/APol complexes at the end of a trapping experiment (12). The latter phenomenon, which is reversible, is probably due to the poor dispersive power of APols: the presence of free polymer favors the formation of MP monomers, while its removal shifts the equilibrium towards protein auto-association. Although few proteins have been tested yet, it seems that freezing MP/APol complexes is not detrimental, while lyophilizing them can be (ref. 9, and unpublished data).
Complexation by A8-35 will, as a rule, stabilize biochemically a MP as compared to detergent solutions (Fig. 3c; see e.g. refs. 1, 5, 6, 9, 10, 18, 20, and refs. therein). The underlying mechanisms are multiple (cf. Table 1 in Applications; for discussions, see refs. 1, 5, 10).
The functionality of APol-trapped MPs is generally preserved (see e.g. refs. 1, 9, 21, 22), as is their ability to bind ligands (reviewed in ref. 16). However, MPs whose functional cycle involves large rearrangements of the surface of their transmembrane region, like the sarcoplasmic calcium ATPase, may see their activity reversibly slowed down or blocked, presumably because the adsorbed polymer damps such transconformations (for a discussion, see refs. 1, 10). The latter phenomenon ('Gulliver effect') probably contributes to slowing down MP denaturation (10).