AMPHIPOLS: Trapping membrane proteins with A8-35
Although some exceptions have been encountered and provide interesting precedents (reviewed in ref. Popot et al. (2003)), A8-35 as a rule will not extract by itself a MP from its native environment. The standard procedure is to solubilize the target protein in a classical detergent and then to transfer it, whether before or after purification, to the polymer (Fig. 3a). This is achieved by forming first a tertiary complex, which happens spontaneously in solutions containing both types of surfactants (see refs. Tribet et al. (2009), Zoonens et al. (2007) and refs. therein). The ratio of polymer to protein to be used varies depending on the molecular mass of the protein and the extension of its transmembrane surface. For small, deeply membrane inserted proteins like BR or tOmpA, a good trapping ratio is typically around 4-5 g A8-35 per g protein (Gohon et al. (2008), Zoonens et al. (2005)), while for a very large complex with extended extra membrane domains like cytochrome bc1 , it can drop to 1.5 g per g (unpublished data). The protein will bind, ultimately, only part of the polymer added. The detergent is then removed by any convenient means, most often by adsorption onto polystyrene beads (see e.g. refs. Gohon et al. (2008), Zoonens et al. (2005), Charvolin et al. (2009) ).
An alternative way of forming a MP/A8-35 complex is to start from the protein in a denaturing environment, typically urea for β-barrel MPs and SDS for α-helical ones, and to (re)fold the protein by adding the APol and removing the denaturant (
Pocanschi et al. (2006),
Dahmane et al. (2009)
; see Folding and Immobilization).
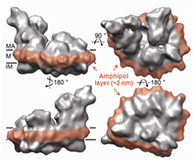
In the final, detergent-free complexes, the polymer associates specifically with the transmembrane surface of the protein ( Zoonens et al. (2005), Catoire et al. (2010, 2009), Althoff et al. (2011)) (Figs. 3b,c), where it forms a compact, relatively thin layer, 1.5 2 nm thick (Gohon et al. (2008), Althoff et al. (2011) ) (Fig. 3c). For various technical reasons, measuring precisely the amount of protein-bound APol is difficult, particularly for small proteins (see refs. Gohon et al. (2008), Zoonens et al. (2007)). Estimates vary from ~2 g per g protein for a small, mainly transmembrane protein like BR (Gohon et al. (2008)) to ~0.11 g per g for a large complex like cytochrome bc1 (Popot et al. (2011)). In most experiments, such as SAXS, SANS, or AUC ones, MP/A8-35 complexes behave like globular proteins (Gohon et al. (2008)). Upon SEC, however, they seem to migrate as though they were somewhat bigger than they actually are (Gohon et al. (2008), Zoonens et al. (2005)), most likely due to repulsive electrostatic interactions with the chromatographic support (Sharma et al. (2012)).
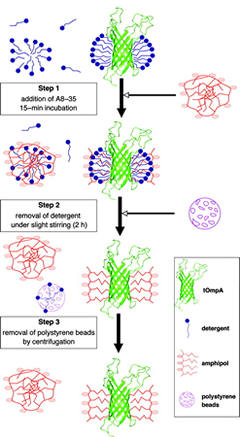
Figure 3a. An example of trapping procedure, in this case using BioBeads (from ref. Zoonens et al. (2005)).
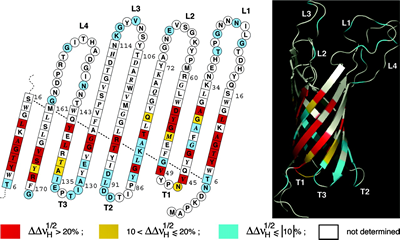
Figure 3b. (above) A map of contacts between A8-35 alkyl chains and the surface of the transmembrane domain of the outer membrane protein OmpA from Escherichia coli. Residues are color-coded depending on whether dipolar line broadening upon substituting a hydrogenated for a deuterated APol is strong (red), weak (yellow), or undetectable (blue). Residues giving rise to no assigned line are white. The data are plotted on a topology sketch and on a ribbon representation of the 3D structure (PDB 1G90) (from ref. Zoonens et al. (2005)).